Lipid membranes play a key role in living systems by providing a structural barrier that separates cellular compartments. Bilayer fluidity in the lateral plane is a key property of lipid membranes, that allows the membrane to have sufficient flexibility to accommodate dynamic stresses, shape changes and rearrangements accompanying the cellular lifecycle.*
In the article “Carbon nanotube porin diffusion in mixed composition supported lipid bilayers” Kylee Sullivan, Yuliang Zhang, Joseph Lopez, Mary Lowe and Aleksandr Noy describe how they used high-speed atomic force microscopy (HS-AFM) and all-atom molecular dynamics (MD) simulations to study the behavior of CNTPs in a mixed lipid membrane consisting of DOPC lipid with a variable percentage of DMPC lipid added to it. HS-AFM data reveal that the CNTPs undergo diffusive motion in the bilayer plane.*
Motion trajectories extracted from the HS-AFM movies indicate that CNTPs exhibit diffusion coefficient values broadly similar to values reported for membrane proteins in supported lipid bilayers. The data also indicate that increasing the percentage of DMPC leads to a marked slowing of CNTP diffusion. MD simulations reveal a CNTP-lipid assembly that diffuses in the membrane and show trends that are consistent with the experimental observations. *
The above-mentioned study confirms that CNTPs mimic the major features of the diffusive movement of biological pores in lipid membranes and shows how the increase in bilayer viscosity leads to a corresponding slowdown in protein motion. It should be possible to extend this approach to studies of other membrane protein dynamics in supported lipid bilayers. The authors note that those studies, however, will need to be mindful of the challenge of unambiguous visualization of the membrane components, especially in systems that incorporate smaller proteins, such as antimicrobial peptides. Another challenge that could complicate these studies would be microscopic phase separation of the lipid matrix that could lead to complicated pore dynamics in the membrane. *
NanoWorld Ultra-Short AFM cantilevers with high-density carbon/diamond-like carbon (HDC/DLC) AFM tips of the USC-F1.2-k0.15 type were used for the high-speed atomic force microscopy described in the article. *

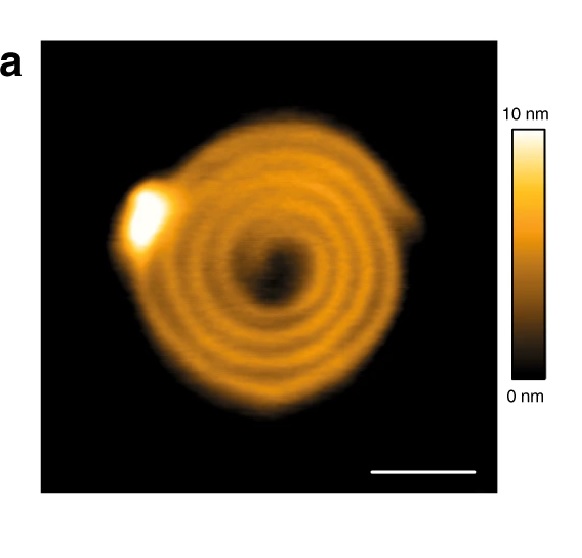
CNTP motion in supported lipid bilayers. (a) Representative frames (with times in seconds indicated on each image) from an HS-AFM movie showing a CNTP diffusing in a supported lipid bilayer with 80:20 DOPC-DMPC ratio (see also Supplementary Movie 2). (b) A representative trajectory for CNTP diffusion in the bilayer. The time step between each datapoint is 0.5 s.
Please refer to the full article cited below for the full figure.
*Kylee Sullivan, Yuliang Zhang, Joseph Lopez, Mary Lowe and Aleksandr Noy
Carbon nanotube porin diffusion in mixed composition supported lipid bilayers
Nature Scientific Reports volume 10, Article number: 11908 (2020)
DOI: https://doi.org/10.1038/s41598-020-68059-2
Please follow this external link to read the full article: https://rdcu.be/b69wj
Open Access : The article “Carbon nanotube porin diffusion in mixed composition supported lipid bilayers” by Kylee Sullivan, Yuliang Zhang, Joseph Lopez, Mary Lowe and Aleksandr Noy is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit https://creativecommons.org/licenses/by/4.0/.