The ongoing coronavirus disease 2019 (COVID-19) pandemic caused by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) represents a serious threat to the health of millions of people. Respiratory viruses such as SARS-CoV-2 can be transmitted via airborne and fomite routes. The latter requires virion adsorption at abiotic surfaces and most likely involves the SARS-CoV-2 spike protein subunit 1 (S1), which is the outermost point of its envelope. Understanding S1 spike protein interaction with fomite surfaces thus represents an important milestone on the road to fighting the spread of COVID-19.*
In the article “Adsorption of SARS-CoV-2 Spike Protein S1 at Oxide Surfaces Studied by High-Speed Atomic Force Microscopy “ Yang Xin, Guido Grundmeier and Adrian Keller describe how high-speed atomic force microscopy (HS-AFM) is used to monitor the adsorption of the SARS-CoV-2 spike protein S1 at Al2O3(0001) and TiO2(100) surfaces in situ. *
Figure 2 from Yang Xin et al Adsorption of SARS-CoV-2 Spike Protein S1 at Oxide Surfaces Studied by High-Speed Atomic Force Microscopy HS-AFM images (1 × 1 μm2) of SARS-CoV-2 spike protein S1 in 10 mM Tris (pH 7.5) adsorbed to a) an Al2O3(0001) and b) a TiO2(100) surface recorded at different time points as indicated. Height scales are 5 nm for the clean substrate surfaces at 0 s and 12 nm for the protein covered surfaces at later time points. Below the HS-AFM images, the corresponding height distribution functions are depicted. The vertical lines in the plots represent the height thresholds applied in the statistical analyses.
*Yang Xin, Guido Grundmeier, Adrian Keller Adsorption of SARS-CoV-2 Spike Protein S1 at Oxide Surfaces Studied by High-Speed Atomic Force Microscopy
Advanced NanoBioMed Research, Volume 1, Issue 2, February 2021, 2000024
DOI: https://doi.org/10.1002/anbr.202000024
Open Access : The article “Adsorption of SARS-CoV-2 Spike Protein S1 at Oxide Surfaces Studied by High-Speed Atomic Force Microscopy” by Yang Xin, Guido Grundmeier and Adrian Keller is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit https://creativecommons.org/licenses/by/4.0/.
Courtesy of Prof. Noriyuki Kodera, nanoLSI, Kanazawa University, Japan we could upload two new images and a very nice High Speed Atomic Force Microscopy (HS-AFM) video of Actin filaments on Mica with APTES to the image gallery and the video gallery on https://www.highspeedscanning.com/.
NanoWorld USC-F1.2-k0.15 Ultra-Short AFM cantilevers [C = 0.15 N/m; fo = 1200 kHz] were used for the high speed imaging.
HS-AFM Images of actin filaments on Mica with APTES. Buffer:100 mM KCl, 2 mM MgCl2, 1 mM EGTA, 20 mM Imidazole-HCl, pH7.6 (a) 250×250 nm2 and (b) 400×400 nm2. Images courtesy of Prof. Kodera, nanoLSI, Kanazawa University, Japan. NanoWorld USC-F1.2-k0.15 AFM probes were used for the High Speed Atomic Force Microscopy
The HighSpeedScanning webpage is dedicated to presenting information about ultra high frequency AFM probe solutions for High Speed AFM ranging from already commercialized AFM probes such as the ArrowTM UHF and NanoWorld Ultra-Short Cantilever (USC) series to AFM probes that are still under development.
Additionally to the High-Speed AFM images and videos researchers worldwide kindly have provided us with so that we can share them with the whole High Speed AFM community you will also find a list of links and references dedicated to High-Speed AFM on https://www.highspeedscanning.com/high-speed-scanning.html
We are aware that these lists are far from complete and we are constantly working on keepting them up to date. If your research institute or company is working with High Speed AFM (HS-AFM) using our AFM probes or if you have published articles with images that were achieved with our High Speed AFM probes annd you find that are missing from our list then please let us know via email: info@highspeedscanning.com if you would like us to add them to the list of references .
Membrane proteins (MPs) reside in the plasma membrane and perform various biological processes including ion transport, substrate transport, and signal transduction.*
Function-related conformational changes in membrane proteins occur in times scales ranging from nanoseconds to seconds.*
Obtaining time-resolved dynamic information of MPs in their membrane environment is still a major challenge.*
Although High Speed Atomic Force Microscopy (HS-AFM) images label-free samples such as DNA, soluble proteins, MPs, and intrinsically disordered proteins at ~1n~m lateral, ~0.1 nm vertical and ~100 ms temporal solution in aqueous environment and at ambient temperature and pressure, its temporal resolution is too slow to characterize many dynamic biological processes.*
In order to overcome this limitation Raghavendar Reddy Sanganna Gari, Joel José Montalvo-Acosta, George R. Heath, Yining Jiang, Xiaolong Gao, Crina M. Nimigean, Christophe Chipot and Simon Scheuring in their article Correlation of membrane protein conformational and functional dynamics use High Speed Atomic Force Microscopy Height Spectroscopy ( HS-AFM-HS) to characterize the microsecond timescale conformational changes of an integral-MP model system, i.e., the outer membrane protein G (OmpG) in a membrane environment.*
The positioning of the AFM tip is guided by HS-AFM imaging immediately before HS-AFM-HS-operation.*
NanoWorld Ultra-Short Cantilevers (USC) of the USC-F1.2-k0.15 type were used for the HS-AFM and HS-AFM-HS presented in the article.*
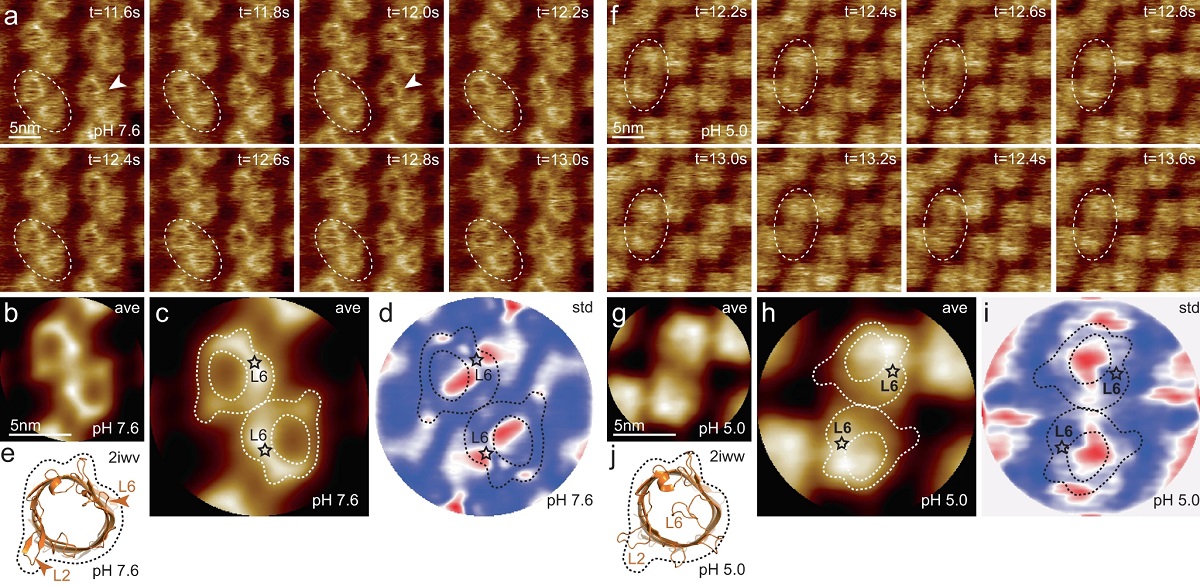
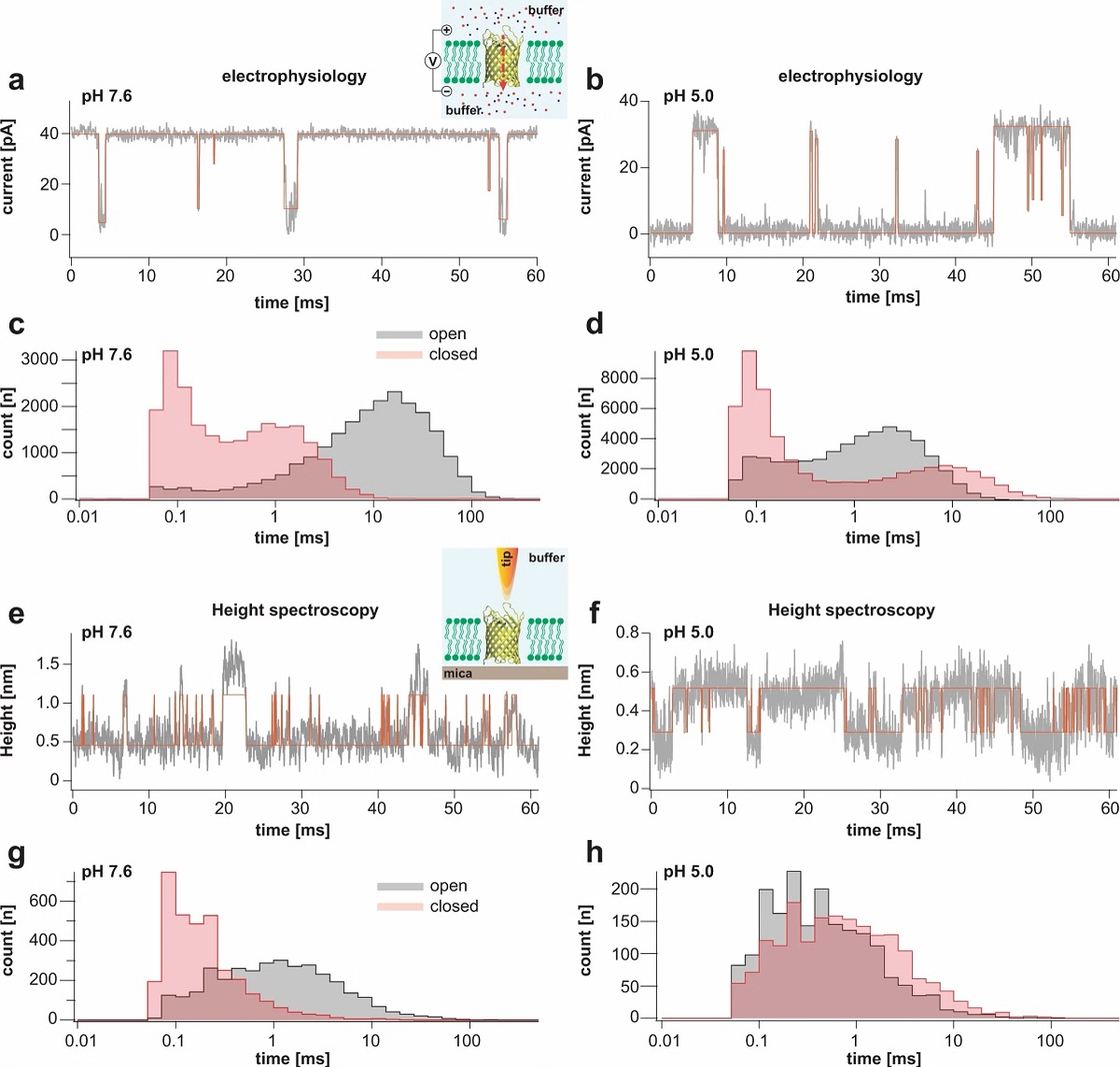
Figure 1 from “Correlation of membrane protein conformational and functional dynamics” by R.R. Sanganna Gari et al: HS-AFM imaging of OmpG in lipid bilayers at pH 7.6 and pH 5.0. a OmpG at pH 7.6 (Supplementary movie 1, left; frame rate: 200 ms per frame). A OmpG dimer is highlighted with dashed outline in all frames. Arrowheads in t = 11.6 s: Loop-6 fluctuating over the lumen. Arrowhead in t = 12.0 s: Fully open state. b Correlation average (n = 2752) of the HS-AFM movie frames (344 frames recorded over 68.8 s, full color scale: 0.0 nm < height < 1.25 nm, where the membrane level was set to 0.0 nm). c Correlation average of OmpG dimers. The topography outline (based on the molecular structure in 1e), serves as a visual guide to locate loop-6 and loop-2 in the topography and is highlighted by the dashed outline (the position of loop-6 is indicated by the asterisk based on its location in the structure (e)). Inner dashed outline show barrel lumen. d Standard deviation (std) map (n = 2752) from the averaging process in (b) (full color scale from blue to red: 0.05 nm < std < 0.19 nm) and topography outlines as in (c). e X-ray structure (PDB 2iwv) of the open OmpG conformation. Loop-6 (arrowhead L6) stands out of the image plane towards the viewer. Loop-2 (L2) forms a beta strand pointing away from the β-barrel, well detected by HS-AFM in the open state (b). f OmpG at pH 5.0 (Supplementary movie 1, right; frame rate: 200 ms per frame). A OmpG dimer is highlighted with dashed outline in all frames. g Correlation average (n = 2472) of the HS-AFM movie frames (309 frames recorded over 61.8 s, full color scale: 0.0 nm < height < 0.7 nm, where the membrane level was set to 0.0 nm). h Correlation average of OmpG dimers. For comparison, the topography outline of the open state (e) is shown (the position of loop-6 is indicated by the asterisk). i Standard deviation (std) map (n = 2472) from the averaging process in (g) (full color scale from blue to red: 0.04 nm < std < 0.07 nm) and topography outlines as in (h). j X-ray structure (PDB 2iww) of the closed OmpG conformation shown in the same orientation as in (e). Loop-6 (L6) folds over the β-barrel lumen in a lid-like manner. Loop-2 (L2) does not form a β-strand in the closed state, in agreement with absence of topography in this region in (h). Black dashed line: outline based on (e) for comparison.Figure 2 from “Correlation of membrane protein conformational and functional dynamics” by R.R. Sanganna Gari et al: Single channel electrophysiology and HS-AFM height spectroscopy recordings of OmpG in lipid bilayers. Representative 60-ms segments of OmpG single channel recordings at pH 7.6 (a) and pH 5.0 (b) at +40 mV membrane potential (longer traces in Supplementary Figs. 2 and 3). Cartoon representation of single channel recording experimental setup is shown in inset of (a). OmpG (yellow) in open state (PDB:2IWV) is placed in a lipid bilayer (green) surrounded by buffer (light blue shade) and potassium and chloride ions are shown as red and blue spheres. Red arrow indicates ion flow through OmpG in response to voltage application. Dwell time histograms of open and closed states at pH 7.6 (c) and pH 5.0 (d) from single-channel recordings (see Supplementary Table 1). Representative 60-ms segments of OmpG HS-AFM-HS recordings at pH 7.6 (e) and pH 5.0 (f) (longer traces in Supplementary Fig. 4). Cartoon representation of HS-AFM height spectroscopy experimental setup is shown in inset of (e). An oscillating AFM tip (orange) detects conformational changes of loop motion. Dwell time histograms of open and closed states at pH 7.6 (g) and pH 5.0 (h) from HS-AFM-HS recordings (Supplementary Table 2). In HS-AFM-HS the low state represents the open state, where the HS-AFM tip can descend into the β-barrel, and the high state represents the closed state, where loop-6 covers the beta barrel barring access of the HS-AFM tip to the cavity. All current-time and height-time traces were filtered at 20 kHz during analysis. The state dwell-time histograms are shown using log binning for better visualization of the components49. Red traces in (a) and (b) represent idealized current-time traces using clampfit software. Red traces in (e) and (f) represent idealized height-time traces using the STaSI algorithm (see Methods).
*Raghavendar Reddy Sanganna Gari, Joel José Montalvo-Acosta, George R. Heath, Yining Jiang, Xiaolong Gao, Crina M. Nimigean, Christophe Chipot and Simon Scheuring Correlation of membrane protein conformational and functional dynamics Nature Communications volume 12, Article number: 4363 (2021) DOI: https://doi.org/10.1038/s41467-021-24660-1
Open Access : The article “Correlation of membrane protein conformational and functional dynamics” by Raghavendar Reddy Sanganna Gari, Joel José Montalvo-Acosta, George R. Heath, Yining Jiang, Xiaolong Gao, Crina M. Nimigean, Christophe Chipot and Simon Scheuring is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this license, visit https://creativecommons.org/licenses/by/4.0/.